
Organizzazione gerarchica dei sistemi motori e riflessi spinali

Riflessi Spinali e sistemi motori
Sistemi motori
I sistemi motori sono in grado di eseguire ad elevata velocità e con grande accuratezza diversi atti motori, sia riflessi che ritmici, che volontari, questo poiché nella loro organizzazione funzionale sono presenti due proprietà:
- l’analisi delle informazioni sensoriali e dei comandi da convogliare ai motoneuroni e ai muscoli è operata da aree interconnesse tra di loro in modo gerarchico, situate in:
- Midollo spinale
- Tronco encefalico
- Proencefalo
- le informazioni sensoriali relative al movimento vengono analizzate a livello di sistemi diversi che operano in parallelo.
Il midollo spinale è il livello più basso di questa organizzazione gerarchica:
- contiene circuiti neuronali che mediano vari riflessi e movimenti automatici di tipo ritmico quali il grattarsi o la locomozione.
- il circuito neuronale nel midollo spinale può essere:
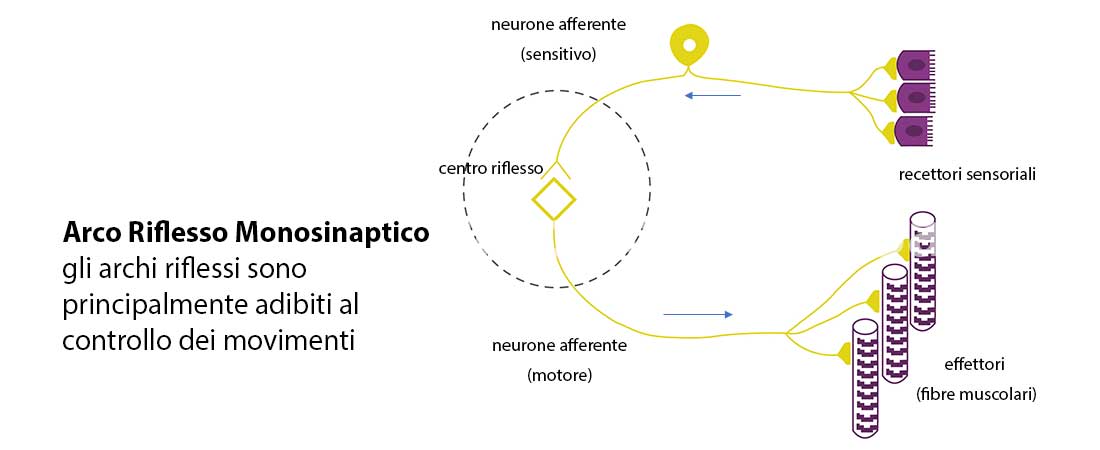
- Monosinaptico: è il circuito più semplice, composto solo da un neurone sensitivo primario e da un motoneurone.
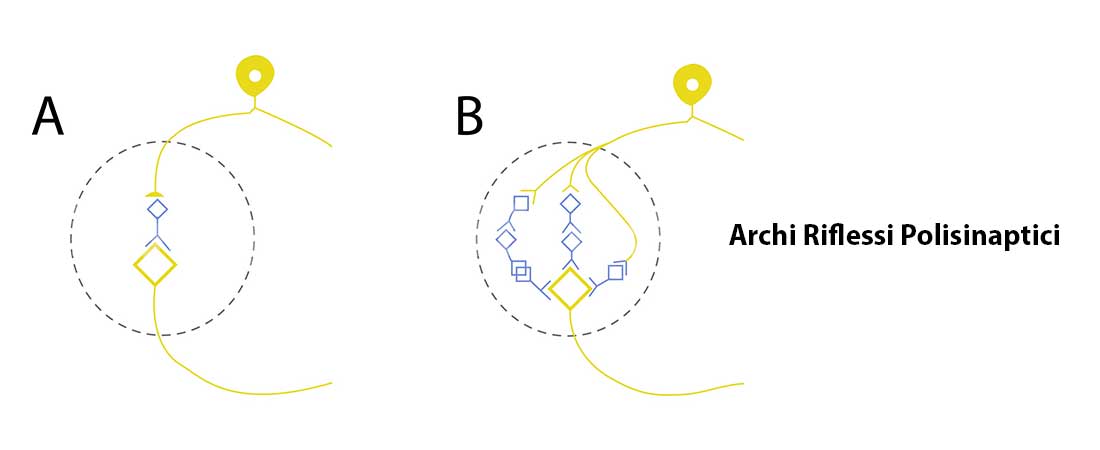
- Polisinaptici: mediano la maggior parte dei riflessi, possiedono uno o più neuroni interposti (interneuroni) tra il neurone sensitivo primario e il motoneurone.
Gli interneuroni e i motoneuroni ricevono anche connessioni da assoni dei centri superiori che hanno la capacità di modificare e coordinare le risposte riflesse agli stimoli periferici (ad esempio, se viene flessa un’articolazione, viene attivato un motoneurone, e l’interneurone che ad esso è connesso inibisce il muscolo antagonista estensore).
Il midollo spinale è un’importante struttura sensitiva e motoria.
Nella parte più ventrale della sostanza grigia i motoneuroni α innervano le fibre muscolari scheletriche mentre i motoneuroni γ innervano i fusi neuromuscolari.
I riflessi spinali possono essere di tipo:
- Somatomotorio: il prodotto finale dell’informazione sensoriale determina una risposta muscolare scheletrica.
- Viscerale: il risultato è una risposta che coinvolge cellule muscolari lisce o ghiandole.
In condizioni normali, le attività riflesse spinali non sono autonome, ma sottoposte al controllo continuo di strutture sovraspinali:
- Tronco dell’encefalo
- Corteccia motoria
Il controllo dei centri superiori è esercitato attraverso vie discendenti che terminano a tutti i livelli della sostanza grigia midollare:
- comandi motori dei centri superiori agiscono solo in piccola parte direttamente sui motoneuroni;
- generalmente, l’attività delle vie discendenti agisce sugli stessi circuiti utilizzati dai riflessi spinali;
- permette l’invio di comandi motori relativamente semplificati.
Per quanto concerne l’organizzazione generale anatomica del midollo spinale, la sostanza grigia midollare può essere suddivisa in:
- versante sensoriale
- versante motorio
- zona di integrazione
In ciascuna metà del midollo si distinguono tre componenti anatomiche principali:
- Corno dorsale: punto di ingresso delle informazioni somatosensitive, che si interrompono a livello del midollo; in esse vi sono localizzati anche i neuroni che trasmettono ai centri superiori attraverso le colonne dorsali o il sistema antero-laterale.
- Zona intermedia: interfaccia tra il versante sensitivo e quello motorio; attraverso questa passa il flusso delle informazioni afferenti dirette verso l’uscita motoria, contiene diversi tipi di cellule nervose, sia eccitatorie che inibitorie.
- Corno anteriore : sede dei motoneuroni.
Il livello successivo di organizzazione gerarchica è il tronco dell’encefalo:
- è costituito da due sistemi di neuroni (laterale e mediale) che ricevono afferenze dalla corteccia cerebrale e da nuclei sottocorticali ed infine proiettano al midollo spinale.
I sistemi discendenti mediali sono implicati nel controllo della postura e integrano informazioni visive, vestibolari e somatosensitive; i sistemi discendenti laterali controllano in misura maggiore i muscoli distali degli arti importanti per i movimenti finalizzati, specialmente quelli della mano e del braccio.
La corteccia costituisce il livello più elevato dell’organizzazione gerarchica del controllo motorio:
corteccia motoria primaria (area F1) e aree premotorie che proiettano direttamente al midollo spinale attraverso il tratto corticospinale coordinando e pianificando delle sequenze motorie complesse.
Altre due formazioni cerebrali partecipano alla pianificazione e all’esecuzione del movimento, il cervelletto e i nuclei della base: i nuclei della base motivano l’azione e fanno scegliere i piani comportamentali, il cervelletto seleziona le caratteristiche temporali e coordina i movimenti in via di svolgimento.
Il sistema motorio periferico è costituito da:
- Unità motorie: complesso del motoneurone e delle fibre muscolari da esso innervate. Le tre unità motorie sono:
- unità FF: motoneuroni grandi, veloci e potenti, ma rapidamente soggette ad affaticamento
- unità SS: motoneuroni piccoli, lente, bassi livelli di forza, resistenti alla fatica
- unità FR: caratteristiche intermedie, ovvero abbastanza veloci e resistenti alla fatica.)
Così come esistono modalità sensoriali diverse, si distinguono diverse categorie di movimenti:
- Riflessi
- Ritmici
- Volontari
Attività motorie basate sui riflessi
I riflessi sono schemi coordinati involontari di contrazioni e rilasciamenti muscolari prodotti da stimoli periferici. L’esame dei riflessi spinali viene utilizzato in clinica per accertare l’integrità delle vie afferenti ed efferenti e per determinare il livello delle lesioni.
Dobbiamo comunque tener presente che esiste un’adattabilità dei riflessi, nonostante tutti i riflessi siano normalmente stereotipati, l’ampiezza del riflesso è aumentata o diminuita secondo necessità comportamentali ad esempio, aumentando il tono muscolare in seguito al potenziamento dell’ampiezza del riflesso patellare da stiramento.
Alcuni adattamenti dei riflessi sono immediati, mentre altri richiedono maggior tempo, necessitano di apprendimento e plasticità sinaitiche, ad esempio un nuovo paio di occhiali da vista che ingrandiscono o rimpiccioliscono l’immagine variano il riflesso vestibolo-oculare.
Attività motorie ritmiche
I movimenti ritmici ripetitivi comprendono:
- Masticazione
- Deglutizione
- Grattamento
- Contrazioni alternate dei flessori e degli estensori di entrambi i lati del corpo (contrazioni cicliche posturali)
I circuiti preposti a queste attività motorie ritmiche ripetitive si trovano in:
- Midollo spinale
- Tronco encefalico (per i muscoli del capo e della faccia).
Movimenti volontari
Differentemente dai riflessi, i movimenti volontari sono diretti al raggiungimento di uno scopo specifico. Il sistema nervoso impara a correggere le perturbazioni esterne in due modi:
- controllo a feedback: utilizza i segnali sensoriali che riceve normalmente per agire direttamente sull’arto che esegue il movimento e correggere nell’immediato il movimento.
- controllo a feedforward: il sistema nervoso utilizza informazioni di una o più modalità diverse (es vista, udito o tatto) per rilevare le perturbazioni cui sta per andare incontro il movimento, ed attuare in maniera anticipatoria delle strategie, sulla base dell’esperienza pregressa.
Riflessi
I riflessi sono movimenti stereotipati evocati dall’attivazione dei recettori cutanei articolari e muscolari.
I riflessi hanno un importante ruolo funzionale nella caratterizzazione dell’attività motoria: si ritiene che i riflessi siano integrati da comandi motori generati dal SNC, al fine di produrre movimenti che possano adattarsi a condizioni ambientali diverse.
I circuiti che mediano le risposte motorie si trovano direttamente nel midollo spinale attivati indipendentemente dal fenomeno percettivo; i centri sovraspinali svolgono un importante ruolo funzionale nella modulazione e nell’adattamento dei riflessi spinali, fino a produrre movimenti di segno opposto in caso di necessità.
Riflessi muscolari
I riflessi muscolari scheletrici sono coinvolti in quasi tutte le nostre azioni.
I recettori che avvertono le variazioni di posizione delle articolazioni, lo stiramento e la tensione muscolare inviano le informazione al SNC, che a sua volta risponde in due modi:
- Se la contrazione muscolare è la risposta appropriata, il SNC attiva i motoneuroni diretti alle fibre muscolari;
- Se, invece, la risposta appropriata richiede che i muscoli si rilascino, l’informazione sensoriale attiva degli interneuroni inibitori del SNA che, a loro volta, inibiscono i neuroni motori che controllano il muscolo.
Gli elementi costitutivi dei riflessi muscolari scheletrici sono:
- i recettori sensoriali, detti propriocettori, sono localizzati all’interno dei muscoli scheletrici, nelle capsule articolari e nei legamenti. Rilevano la posizione dei vari segmenti del nostro corpo nello spazio, i nostri movimenti e lo sforzo che esercitiamo;
- i neuroni sensoriali che conducono l’informazione dai propriocettori al SNC (il sistema nervoso centrale integra i segnali di ingresso attraverso circuiti modulatori costituiti da interneuroni sia eccitatori che inibitori);
- i motoneuroni somatici trasportano i segnali in uscita. I motoneuroni somatici, che innervano le fibre contrattili del muscolo scheletrico, sono detti α-motoneuroni;
- gli effettori sono le fibre contrattili del muscolo scheletrico che ricevono i potenziali d’azione dei motoneuroni alfa determinando la contrazione muscolare.
A livello del muscolo scheletrico si trovano tre tipi di propriocettori:
- i fusi muscolari,
- gli organi tendinei del Golgi
- i meccanorecettori articolari
Quest’ultimi, i meccanorecettori articolari, si trovano nei legamenti di ogni articolazione e vengono stimolati da distorsioni meccaniche.
I fusi neuromuscolari sono recettori di stiramento, cioè inviano al midollo spinale e all’encefalo informazione relative alla lunghezza muscolare. I fusi sono strutture piccole, di forma allungata e disposte in parallelo con le fibre muscolari.
Ogni fuso è costituito da una capsula di tessuto connettivo che avvolge un gruppo di piccole fibre muscolari dette fibre intrafusali.
Quest’ultime sono fibre muscolari modificate, per cui le loro estremità sono contrattili mentre la parte centrale è priva di miofibrille. Le estremità delle fibre sono innervate da motoneuroni gamma. La porzione centrale, invece, è avvolta da terminazioni di neuroni sensoriali che sono sensibili allo stiramento. Questi neuroni sensoriali proiettano al midollo spinale e fanno sinapsi direttamente con i motoneuroni alfa che innervano lo stesso muscolo entro cui si trovano i fusi da cui viene l’afferenza.
Quindi, quando un muscolo è alla lunghezza di riposo, la porzione centrale del fuso è sufficientemente stirata da attivare le fibre sensoriali. Per questo i neuroni sensoriali afferenti dai fusi sono tonicamente attivi e inviano un flusso continuo di potenziali d’azione al SNC. A causa di questa attività tonica, anche un muscolo a riposo mantiene sempre un certo livello di tensione, che viene detta tono muscolare.
Quindi ogni movimento che aumenta la lunghezza muscolare stira anche i fusi e causa un aumento della frequenza di scarica della fibra sensoriale. Questo innesca la contrazione riflessa del muscolo, facendo sì che la lunghezza del muscolo rimanga costante ed evitando il danno determinato da un eccessivo stiramento. Questo riflesso, in cui l’allungamento del muscolo innesca una risposta contrattile è detto riflesso da stiramento o riflesso miotatico.
Riflesso da stiramento
Il riflesso più importante, e senza dubbio il più studiato, è il riflesso da stiramento o riflesso miotatico:
prevede la contrazione di un muscolo provocata dal suo allungamento, all’interno delle fibre muscolari i fusi neuromuscolari rilevano variazioni di lunghezza, le fibre afferenti dai fusi neuromuscolari stabiliscono connessioni eccitatorie dirette con i motoneuroni α; si ha così oltre alla contrazione del muscolo stirato, anche l’inibizione dei muscoli antagonisti (innervazione reciproca, essa la ritroviamo nei riflessi, ad esempio lo abbiamo nel riflesso flessorio che prevede la retrazione brusca dell’arto, con flessione di tutte e tre le articolazioni, in seguito all’applicazione di stimoli nocivi; è un riflesso polisinaptico in cui l’attivazione muscolare si prolunga oltre la durata dello stimolo.
Nel riflesso flessorio, la flessione dell’arto stimolato si accompagna all’estensione dell’arto controlaterale corrispondente che permette lo scarico del peso corporeo sull’arto contro laterale e il mantenimento dell’equilibrio. Il riflesso di innervazione reciproca si ritrova anche nei movimenti volontari, il rilasciamento del muscolo antagonista nel corso dell’esecuzione dei movimenti volontari ne aumenta la velocità e l’efficienza; essi ricevono segnali dalle vie collaterali degli assoni discendenti dei neuroni della corteccia motrice che innervano direttamente i motoneuroni spinali).
La velocità di segnale a livello di una sinapsi di tale circuito è 0,5-0,9 ms, di conseguenza si deduce che le fibre stabiliscono connessioni dirette con i motoneuroni α, il riflesso è generato da una via monosinaptica.
L’organo tendineo del Golgi
Un secondo tipo di propriocettore muscolare è l’organo tendineo del Golgi.
Questi recettori sono localizzati a livello della giunzione tra i tendini e le fibre muscolari e rispondono principalmente alla tensione sviluppata dal muscolo durante la contrazione isometrica, e causano un riflesso di rilassamento. Questo è l’opposto di quanto succede per i fusi neuromuscolari, che provocano una contrazione fissa.
Gli organi tendinei del Golgi sono costituiti da terminazioni nervose libere, intrecciate tra fibre di collagene all’interno di una capsula di tessuto connettivo. Quando il muscolo si contrae, i tendini si stirano e questa contrazione sottopone a una trazione le fibre collagene dell’apparato del Golgi, determinando una pressione sulle terminazioni sensoriali dei neuroni afferenti causando la loro attivazione.
L’attivazione dell’organo tendineo del Golgi eccita degli interneuroni inibitori nel midollo spinale, che a loro volta inibiscono i motoneuroni alfa che innervano il muscolo, quindi la contrazione muscolare diminuisce o cessa. Nella maggior parte dei casi, questo riflesso rallenta la contrazione muscolare quando la forza di contrazione aumenta.
In altri casi, gli organi tendinei del Golgi prevengono l’eccessiva contrazione che potrebbe danneggiare il muscolo. L’organo tendineo di Golgi codifica in maniera continuativa la forza del muscolo, in caso della sua contrazione.
Bibliografia:
- Neuroanatomia con riferimenti funzionali e clinici di Turlough M. J. Fitzgerald, Gregory Gruener, Estomih Mtui – Elsevier
- Vander, Sherman, Luciano. Fisiologia dell’uomo.
- Cattaneo L., (1989) Anatomia del Sistema nervoso centrale e periferico dell’uomo. Monduzzi Editore, Bologna. ISBN 88-323-0506-2
- Heckmann CJ, Gorassini MA, Bennett DJ (February 2005). “Persistent inward currents in motoneuron dendrites: implications for motor output”. Muscle Nerve 31 (2): 135–56.doi:10.1002/mus.20261. PMID 15736297.